Anatomia comparada
Dentes que contam histórias
O que um dente fóssil revela sobre como o animal caçava? Em paleontologia, o esmalte é uma das peças mais ricas do quebra-cabeça. Pelo formato, pela serrilha, pela espessura e pela curvatura, paleontólogos reconstruem dieta e estratégia de caça com surpreendente precisão. Esta página compara nove predadores e mostra que dentes parecidos no microscópio podem revelar comportamentos completamente diferentes.
Como ler um dente predador
Antes de comparar espécies, vale entender três variáveis que paleontólogos usam para classificar dentes carnívoros. Quase todas as diferenças que veremos a seguir aparecem em alguma combinação dessas três.
1. Formato em corte
Um dente é zifodonte quando é achatado lateralmente, em forma de lâmina, como uma faca. É conidonte quando o corte transversal é redondo, em forma de cone, como um espinho. Lâminas cortam, cones perfuram. A escolha entre as duas formas costuma indicar se o predador atacava presas grandes (lâmina, para fatiar) ou presas escorregadias (cone, para perfurar e segurar).
2. Serrilhas (denticles)
Pequenos dentes nas bordas, como uma faca de pão. Quanto mais finas e regulares, mais especializadas em cortar carne. Carcarodontossaurídeos têm serrilhas tão regulares que o nome do grupo significa "lagarto com dente de tubarão" (de Carcharodon, o gênero do tubarão branco). Espinossaurídeos, em contraste, têm serrilhas reduzidas ou ausentes, sinal claro de dieta diferente.
3. Robustez e raiz
Um dente fino e curvo é eficiente para cortar, mas quebra ao bater em osso. Um dente grosso, com raiz profunda no maxilar, aguenta mordidas de impacto altíssimo. O T-Rex levou esse padrão ao extremo: dentes "engrossados" (incrassados) com raiz quase do mesmo tamanho da coroa, capazes de mordidas com mais de 3,5 toneladas de força sem se partirem.
Escola 1
Os esmagadores de osso
Quando o predador é grande o suficiente para morder vértebras de presas e processar carcaças inteiras, dentes finos como faca não servem: quebrariam. A solução evolutiva foi engrossar tudo. Os tiranossaurídeos, e o T-Rex em particular, são o caso mais extremo dessa estratégia em toda a história dos dinossauros.


O que o dente diz sobre a caça
Os dentes do T-Rex eram incrassados: a base é quase tão larga quanto o comprimento do dente, e o esmalte é mais espesso que o de qualquer outro terópode conhecido. Erickson e colaboradores (1996) e estudos posteriores de elementos finitos mostraram que essa geometria distribui o estresse de uma mordida brutal sem fraturar o dente.
A força da mordida foi estimada por Bates e Falkingham (2012) e Gignac e Erickson (2017) entre 3.500 e 6.400 kg-f, a mais alta de qualquer animal terrestre conhecido, vivo ou extinto. Uma combinação de dentes engrossados com força extrema permitia o que paleontólogos chamam de "puncture-and-pull": morder com força total, perfurando carne e osso, e puxar a cabeça para arrancar pedaços. Marcas de dentes do T-Rex em fósseis de Triceratops e Edmontosaurus mostram exatamente esse padrão.
A estratégia tinha um custo: dentes incrassados são ineficientes para cortar, comparados a lâminas finas. O T-Rex compensava esse problema substituindo dentes ao longo da vida (assim como outros terópodes), em ciclos de cerca de 2 anos por dente, segundo D'Emic et al. (2019).
Escola 2
Os fatiadores de carne
Carcarodontossaurídeos e alossaurídeos seguiram a estratégia oposta à do T-Rex. Em vez de engrossar o dente para suportar mordidas em osso, afinaram para uma especialização brutal: cortar carne em fatias profundas. Essa "escola" inclui os maiores predadores que já existiram, e dominou o topo da cadeia alimentar do meio do Jurássico ao meio do Cretáceo.
Carcharodontosaurus saharicus
"Lagarto com dentes de tubarão", Cretáceo Médio (~99 a 94 Ma), África do Norte

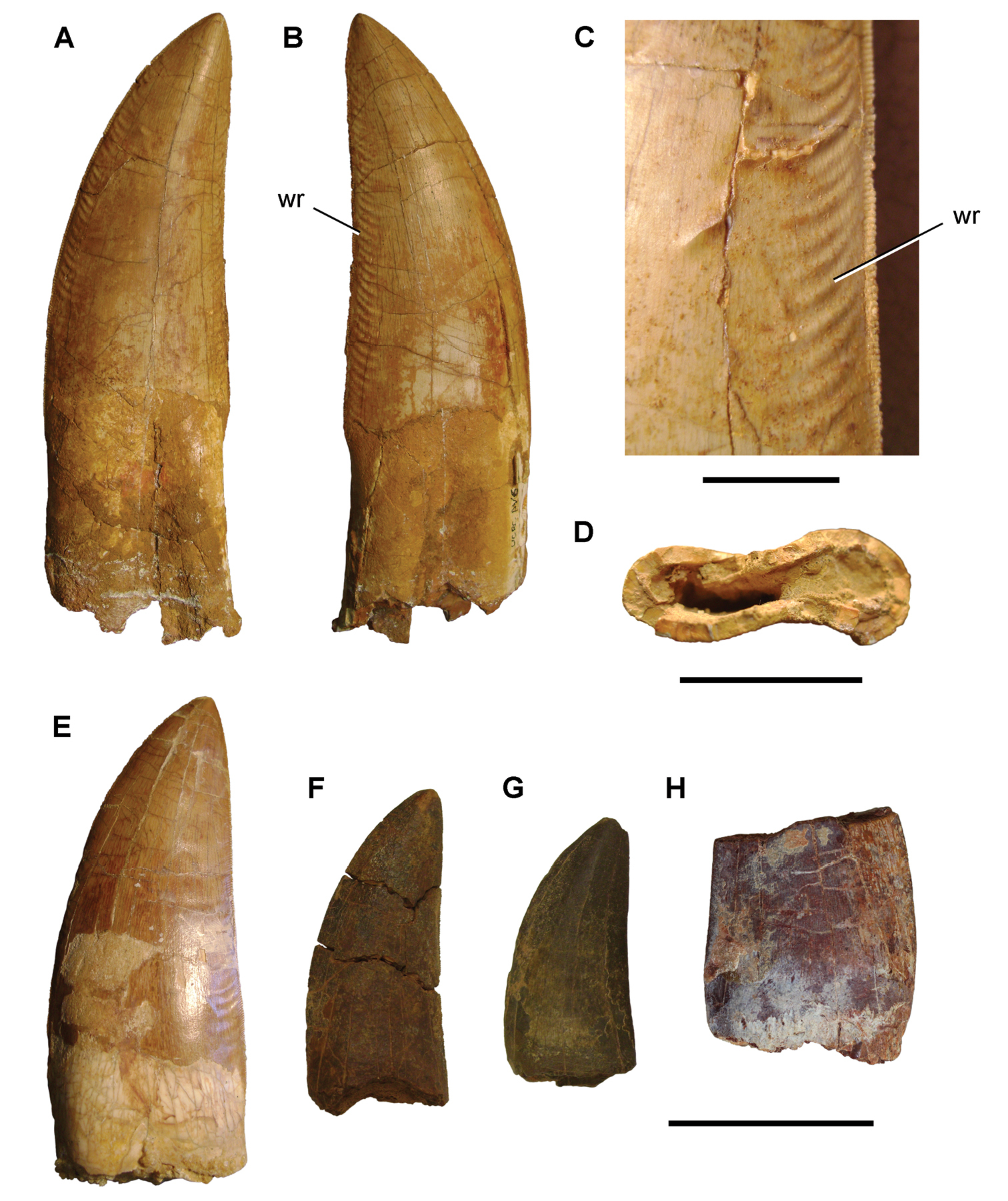
Giganotosaurus carolinii
Cretáceo Superior (~99 a 95 Ma), Patagônia argentina. Primo sul-americano do Carcharodontosaurus.


Allosaurus fragilis
Jurássico Superior (~155 a 145 Ma), América do Norte. Primo mais antigo dos carcarodontossaurídeos.

O que o dente diz sobre a caça
Para Carcharodontosaurus e Giganotosaurus, a interpretação é direta: dentes desenhados para fatiar carne em pedaços profundos, especialmente de saurópodes. Estudos de força de mordida em carcarodontossaurídeos (Bates e Falkingham, 2018) estimam algo em torno de 2.000 a 3.000 kg-f, abaixo do T-Rex, mas suficiente para penetrar a pele espessa de titanossauros. A estratégia dependia do volume de sangue perdido pela presa: cortes profundos e repetidos até a presa entrar em colapso, semelhante ao comportamento descrito hoje em tubarões brancos atacando focas.
O Allosaurus segue o mesmo desenho geral, mas com um crânio mais leve e dentes proporcionalmente menores. Rayfield (2001) propôs a famosa hipótese da "mordida de machado": o crânio do Allosaurus suportava bem cargas verticais, mas mal cargas laterais. A interpretação é que ele usava o crânio como machado, abaixando a cabeça com força do pescoço sobre a presa, em vez de morder e tracionar como o T-Rex.
Em todos os três casos, os dentes serrilhados deixam um padrão característico em ossos: marcas paralelas finas, distintas das marcas pontudas e largas deixadas pelos dentes incrassados do T-Rex. Esse contraste é o que permite identificar o autor de uma mordida em fósseis com mais de 100 milhões de anos.
Escola 3
Os pegadores de peixe
Os espinossaurídeos quebram completamente o padrão dos terópodes carnívoros. Em vez de lâminas serrilhadas, têm dentes cônicos, lisos, sem serrilhas. O resultado parece dente de crocodilo, e não é coincidência: a função é a mesma, prender presas escorregadias e segurá-las até a deglutição.
Spinosaurus aegyptiacus
Cretáceo Médio (~99 a 93 Ma), Norte da África. O maior terópode conhecido, possivelmente semiaquático.


Baryonyx walkeri
Cretáceo Inferior (~125 Ma), Inglaterra. Primo menor e mais antigo do Spinosaurus.

O que o dente diz sobre a caça
Dentes cônicos sem serrilha são péssimos para cortar carne e ótimos para perfurar e segurar. É exatamente o oposto da escola dos fatiadores. O Baryonyx é o caso mais conclusivo: o holotipo descrito por Charig e Milner (1986) preservou no estômago do animal escamas e dentes parcialmente digeridos do peixe Lepidotes, evidência direta de dieta piscívora. Foi o primeiro dinossauro com conteúdo estomacal de peixe encontrado.
Para o Spinosaurus, a evidência é mais indireta mas convergente. Ibrahim e colaboradores (2014, 2020) reanalisaram fósseis e propuseram que o Spinosaurus era o primeiro terópode semiaquático conhecido: cauda em forma de remo, narinas em posição alta, ossos densos para flutuabilidade reduzida. A morfologia dental encaixa nesse cenário. Beevor et al. (2021) estudou a quantidade massiva de dentes de Spinosaurus em jazidas fluviais do Kem Kem e concluiu que o animal passava boa parte da vida na água, alimentando-se de peixes como o pré-histórico Onchopristis, uma raia-serra com dois metros de comprimento.
Resumo da escola: o dente cônico é uma assinatura ecológica. Um terópode com dentes assim quase certamente comia peixe ou outra presa escorregadia. A morfologia conservou-se na linhagem dos espinossaurídeos por mais de 30 milhões de anos.
Escola 4
Os pequenos coadjuvantes
Nem todo predador conta principalmente com os dentes. Os dromeossaurídeos, grupo do Velociraptor e do Deinonychus, têm dentes pequenos, recurvados e serrilhados, parecidos com versões em miniatura dos zifodontes. Mas a arma principal estava no pé: a famosa garra-foice retrátil do segundo dedo. Os dentes vinham depois.


O que o dente diz sobre a caça
Os dromeossaurídeos são o exemplo mais claro de que dente sozinho não conta a história toda. Sua arma principal era a garra retrátil do segundo dedo do pé, que podia chegar a 13 cm em Deinonychus. Fowler et al. (2011) propuseram a hipótese do "raptor prey restraint" (RPR), ou contenção da presa: o predador subia sobre presas menores e usava o peso corporal mais a garra para imobilizá-las, mordendo a presa viva enquanto ela ainda se debatia. Essa hipótese é apoiada por estudos de comportamento de aves de rapina modernas (águias, falcões), que matam suas presas exatamente assim.
Nessa estratégia, os dentes funcionam como ferramentas de processamento, não de caça. Eles arrancam pedaços de carne enquanto o pé e o peso seguram o animal. É bem diferente do T-Rex ou do Carcharodontosaurus, em que a mordida é a própria forma de matar.
O caso famoso do "Fighting Dinosaurs", fóssil mongol descoberto em 1971 com um Velociraptor entrelaçado em um Protoceratops, mostra a garra do raptor afundada na garganta do herbívoro, e os dentes sem registro de uso letal. A coreografia da caça está toda no pé.
Fora da curva
Três casos que quebram a regra
Algumas espécies não se encaixam no espectro principal. Elas mostram que evolução não é linear: o mesmo grupo (terópodes) pode produzir um animal que come peixe, outro que esmaga osso, outro que seguraria presas como um felino e até um terópode que virou herbívoro.
Majungasaurus crenatissimus
Cretáceo Superior (~70 a 66 Ma), Madagascar. O único grande predador da ilha.

Estratégia: morder e segurar
Snively et al. (2011) usaram análise de elementos finitos para reconstruir a mecânica do crânio do Majungasaurus. A conclusão: estratégia parecida com a de felinos modernos, morder e segurar a presa enquanto a musculatura do pescoço fazia o resto. Os dentes mais curtos e robustos suportavam carga sustentada durante a luta com a presa, não a mordida única dos fatiadores.
A presa principal era o Rapetosaurus krausei, titanossauro de cerca de 15 metros. Há marcas de dentes do Majungasaurus em ossos de Rapetosaurus, identificadas por Rogers et al. (2007).
E há um detalhe macabro: marcas de dentes de Majungasaurus em ossos de Majungasaurus, descritas por Rogers, Krause e Curry Rogers (2003) na Nature. É o único caso bem documentado de canibalismo em dinossauros não avianos. Seria predação ativa ou consumo de carcaças? Ainda em debate.
Therizinosaurus, Erlikosaurus, Falcarius
Cretáceo (~125 a 70 Ma), Ásia e América do Norte. Terópodes que viraram herbívoros.
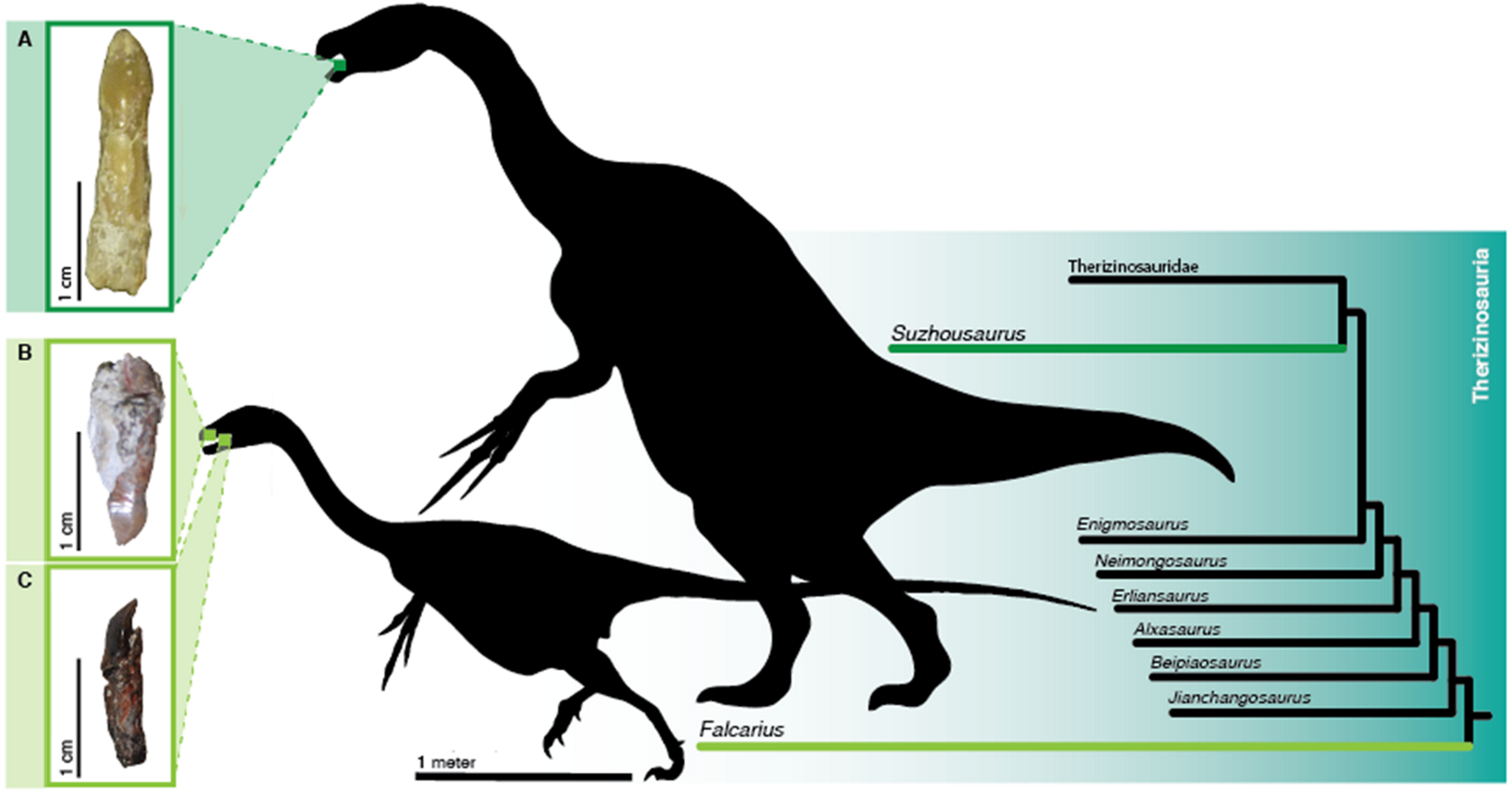

A reviravolta evolutiva
Terizinossaurídeos eram terópodes, o mesmo grupo do T-Rex e do Velociraptor. Filogeneticamente, são parentes próximos das aves e dos dromeossaurídeos. Mas durante o Cretáceo, alguma linhagem fez a transição mais radical possível: deixou de ser carnívora e virou herbívora.
A evidência está nos dentes. Button et al. (2017) analisaram a histologia dos dentes de Falcarius e Suzhousaurus e mostraram que houve mudanças no padrão de incremento dos tecidos compatíveis com dieta vegetal. Os dentes em folha são ineficazes para cortar carne, mas eficientes para esfarelar vegetação. Combinados com bicos anteriores queratinosos (como em aves), pescoços longos e barrigas enormes para fermentação intestinal, configuram uma "máquina de comer planta" disfarçada de carnívoro.
O caso quebra uma intuição simples: terópode não é sinônimo de carnívoro. A linhagem é definida pela anatomia geral (bipedalismo, ossos ocos, etc), não pela dieta. E o dente, mais uma vez, é o sinalizador da mudança.
Carnotaurus sastrei
Cretáceo Superior (~72 a 70 Ma), Patagônia argentina. Abelissaurídeo de cara curta com chifres.

Estratégia: mordida rápida com pescoço potente
Mazzetta e colaboradores (1998, 2004, 2009) modelaram a mecânica do crânio do Carnotaurus e chegaram a um resultado curioso: ele não tinha mordida especialmente forte. A força estimada é menor do que a de um Allosaurus de tamanho similar. Mas o pescoço era enormemente musculoso: as cervicais têm áreas de inserção muscular comparáveis às de touros e bisões.
A interpretação consensual: o Carnotaurus dava mordidas curtas e rápidas em movimento, contando com o pescoço para gerar a força do impacto. Os dentes pequenos eram suficientes para o tipo de caça, e a cara curta diminuía o momento (alavanca) na hora de bater. Faz pensar em um leão correndo lateralmente atrás de uma gazela e dando dentadas em série, ao invés de um T-Rex parando uma presa com força bruta.
Carnotaurus também tinha braços notavelmente curtos e atrofiados, ainda mais que os do T-Rex. A musculatura do pescoço e a velocidade compensavam tudo. É outro lembrete de que dente é só uma peça: para entender como um predador caçava, é preciso olhar o crânio inteiro, o pescoço, e o resto do corpo.
Visão geral: cinco escolas, nove predadores
Cada predador adaptou seus dentes ao tipo de presa, ao tamanho do próprio corpo e ao que o resto da anatomia oferecia. Pequenas diferenças de geometria correspondem a estratégias de caça radicalmente distintas.
| Predador | Tipo de dente | Estratégia de caça | Presa típica |
|---|---|---|---|
| T-Rex | Banana, incrassado, raiz profunda | Esmaga osso (puncture-and-pull) | Hadrossauros, Triceratops |
| Carcharodontosaurus | Lâmina muito achatada, serrilhada | Fatia carne em cortes profundos | Saurópodes (titanossauros) |
| Giganotosaurus | Lâmina, idêntico ao Carcharodontosaurus | Fatia carne, possivelmente em grupo | Saurópodes patagônicos |
| Allosaurus | Lâmina, mais curta e robusta | Mordida de machado (golpe vertical) | Saurópodes jurássicos, estegossauros |
| Spinosaurus | Cônico, liso, sem serrilha | Pesca em rio (semiaquático) | Peixes, raias-serra (Onchopristis) |
| Baryonyx | Cônico, focinho longo (gavial-like) | Pesca em rio (confirmada por estômago) | Peixes (Lepidotes) |
| Velociraptor / Deinonychus | Pequeno recurvado, serrilhado | Garra-foice imobiliza, dente processa | Protoceratops, presas pequenas |
| Majungasaurus | Curto, robusto, base arredondada | Morde e segura (estilo felino) | Rapetosaurus, canibalismo |
| Therizinosaurus | Folha, sem serrilha afiada | Não caça: é herbívoro | Folhas, frutos, talvez insetos |
| Carnotaurus | Pequeno para o tamanho do animal | Mordida rápida com pescoço potente | Presas médias, ataques em movimento |
Conclusão
Um dente fóssil é um documento. Pelo formato, pela serrilha, pela espessura e pela curvatura, paleontólogos reconstroem o que o animal comia, como atacava, e até comportamentos sociais. A diversidade de soluções dentárias entre dinossauros predadores é prova de que o nicho de "predador de topo" foi preenchido por estratégias muito diferentes ao longo do Mesozoico, do esmagador de osso americano ao pescador de rio africano, do canibal insular malgaxe ao terópode que virou herbívoro.
Fontes
- Bates, K. T. & Falkingham, P. L. (2012). Estimating maximum bite performance in Tyrannosaurus rex. Biology Letters, 8(4), 660-664.
- Bates, K. T. & Falkingham, P. L. (2018). The importance of muscle architecture in biomechanical reconstructions of extinct animals. J. Anatomy.
- Beevor, T. et al. (2021). Taphonomic evidence supports an aquatic lifestyle for Spinosaurus. Cretaceous Research, 117, 104627.
- Brownstein, C. D. (2019). New records of theropods from the latest Cretaceous of New Jersey. Royal Society Open Science.
- Button, K., You, H., Kirkland, J. I. & Zanno, L. E. (2017). Incremental growth of therizinosaurian dental tissues. PeerJ, 5:e4129.
- Charig, A. J. & Milner, A. C. (1986). Baryonyx, a remarkable new theropod dinosaur. Nature, 324, 359-361.
- Cuff, A. R. & Rayfield, E. J. (2013). Feeding mechanics in spinosaurid theropods and extant crocodilians. PLOS ONE, 8(5).
- D'Emic, M. D. et al. (2019). Evolution of high tooth replacement rates in theropod dinosaurs. PLOS ONE, 14(11).
- Erickson, G. M. et al. (1996). Bite-force estimation for Tyrannosaurus rex from tooth-marked bones. Nature, 382, 706-708.
- Fowler, D. W. et al. (2011). The predatory ecology of Deinonychus and the origin of flapping in birds. PLOS ONE, 6(12).
- Gignac, P. M. & Erickson, G. M. (2017). The biomechanics behind extreme osteophagy in Tyrannosaurus rex. Scientific Reports, 7, 2012.
- Ibrahim, N. et al. (2014). Semiaquatic adaptations in a giant predatory dinosaur. Science, 345, 1613-1616.
- Ibrahim, N. et al. (2020). Tail-propelled aquatic locomotion in a theropod dinosaur. Nature, 581, 67-70.
- Mazzetta, G. V., Cisilino, A. P., Blanco, R. E. & Calvo, N. (2009). Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei. Journal of Vertebrate Paleontology, 29(3), 822-830.
- Rayfield, E. J. et al. (2001). Cranial design and function in a large theropod dinosaur. Nature, 409, 1033-1037.
- Rogers, R. R., Krause, D. W. & Curry Rogers, K. (2003). Cannibalism in the Madagascan dinosaur Majungatholus atopus. Nature, 422, 515-518.
- Rogers, R. R. et al. (2007). Paleoenvironment and taphonomy of the Anembalemba Member, Maevarano Formation. SVP Memoir 8.
- Snively, E. et al. (2011). Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods. PeerJ.